Evolving Neural Networks through Augmenting Topologies (NEAT)
作者: Kenneth O. Stanley and Risto Miikkulainen (The University of Texas at Austin) 发表时间: 2002 (Based on TR 2001)
1. 引言 (Introduction)
神经进化 (Neuroevolution, NE) 是利用遗传算法来训练神经网络的方法。传统的神经进化方法通常固定神经网络的拓扑结构(即神经元和连接的数量及布局),仅进化连接的权重。
然而,神经网络的拓扑结构(Topology)对其功能也至关重要。一个核心问题是:同时进化拓扑结构和权重是否能比仅进化权重带来优势?
早期关于进化拓扑结构的研究(TWEANNs)结论不一,甚至有观点认为固定拓扑结构(如 ESP 算法)效率更高。本文提出了 NEAT (NeuroEvolution of Augmenting Topologies) 方法,旨在通过正确的方式进化拓扑结构,从而显著提升学习效率。
NEAT 的设计旨在解决进化拓扑结构时的三个主要技术挑战: 1. 交叉 (Crossover):如何在不同的拓扑结构之间进行有意义的交叉?(解决竞争约定问题) 2. 保护创新 (Protecting Innovation):如何防止新产生的结构(往往初始适应度较低)在优化前被淘汰? 3. 最小化结构 (Minimizing Structure):如何确保网络在整个进化过程中保持尽可能简单,从而最小化搜索空间?
2. 背景与挑战 (Background & Challenges)
2.1 竞争约定问题 (The Competing Conventions Problem)
这是神经进化中的一个主要难题。对于同一个神经网络功能,存在无数种不同的表达方式(例如隐藏层神经元的排列顺序不同)。如果两个父代网络功能相同但编码(基因排列)不同,直接交叉往往会产生功能受损的后代。
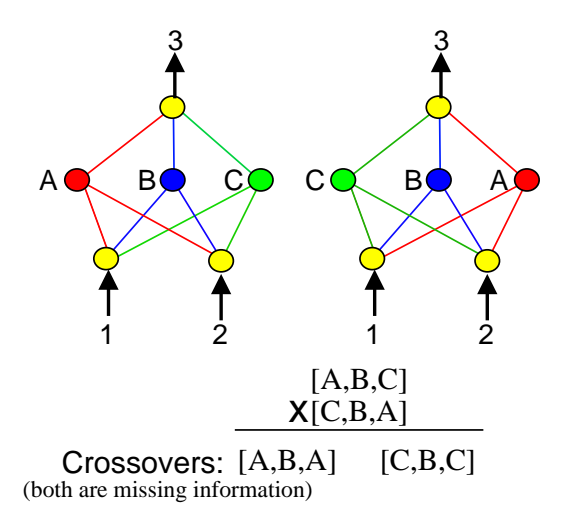 图1:竞争约定问题示意图。两个网络计算相同的函数,但隐藏节点的顺序不同。直接交叉会导致关键信息的丢失(如 [C, B, C] 丢失了 A 的信息)。
图1:竞争约定问题示意图。两个网络计算相同的函数,但隐藏节点的顺序不同。直接交叉会导致关键信息的丢失(如 [C, B, C] 丢失了 A 的信息)。
当网络拓扑结构不同时,这个问题变得更加严重。不知道哪个基因对应哪个结构,交叉就变成了乱序组合。
2.2 保护创新
在 TWEANN 中,通过变异增加新的结构(如新节点或新连接)通常会暂时降低网络的适应度(Fitness)。如果直接与整个种群竞争,这些拥有新结构但尚未优化的个体很快就会死掉。这就像自然界中的新物种需要一个生态位(Niche)来生存一样。
3. NEAT 方法 (The NEAT Method)
NEAT 通过以下三个核心技术解决了上述问题:
3.1 遗传编码 (Genetic Encoding)
NEAT 使用直接编码。每个基因组包含两部分列表: 1. 节点基因 (Node Genes):列出输入、隐藏和输出节点。 2. 连接基因 (Connection Genes):列出连接的起点、终点、权重、是否启用 (Enable bit) 以及最重要的 创新号 (Innovation Number)。
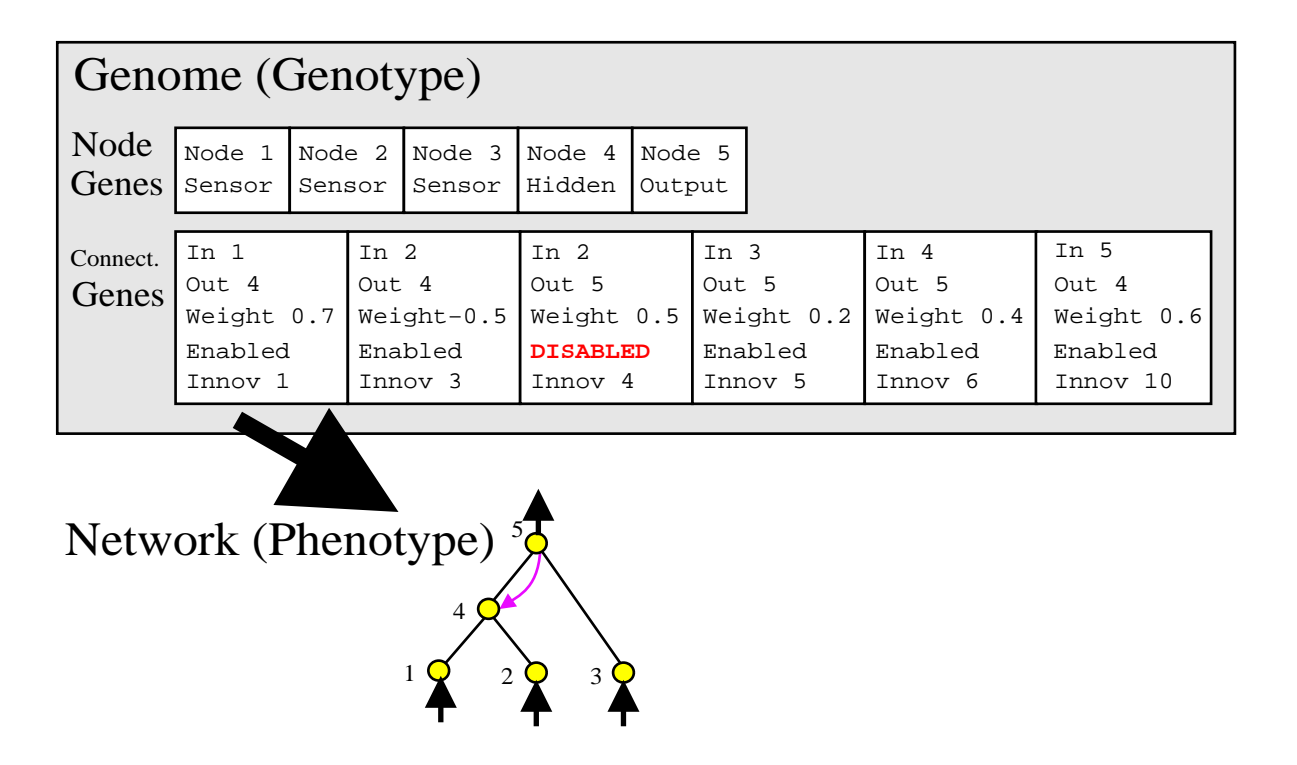 图2:基因型到表现型的映射。基因组(上)通过连接基因直接描述了神经网络(下)。注意第三个基因被禁用(Disabled),因此连接 2->5 在网络中不显示。
图2:基因型到表现型的映射。基因组(上)通过连接基因直接描述了神经网络(下)。注意第三个基因被禁用(Disabled),因此连接 2->5 在网络中不显示。
3.2 历史标记 (Historical Markings) - 解决交叉问题
NEAT 的核心创新在于使用 创新号 (Innovation Number) 来跟踪基因的历史起源。 * 每当通过变异产生一个新的连接基因时,系统会分配一个全局唯一的创新号。 * 继承自同一祖先的基因将拥有相同的创新号。
通过创新号,系统确切地知道不同基因组中的哪些基因是匹配的(同源的),哪些是不匹配的。
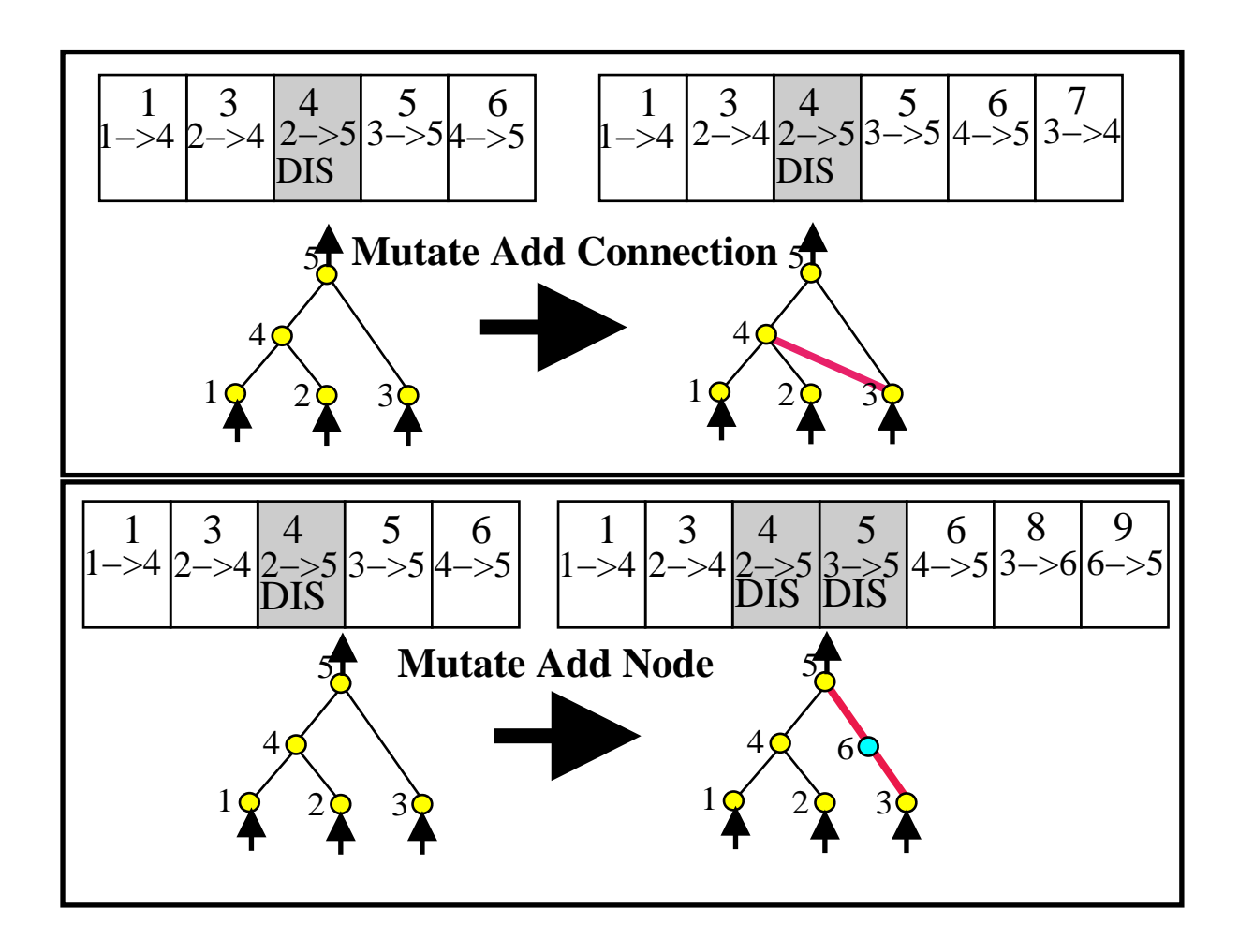 图3:NEAT 的两种结构变异。
1. 添加连接 (Add Connection):在两个节点间增加新连接,赋予新的创新号。
2. 添加节点 (Add Node):在现有连接中间插入新节点。原连接被禁用,产生两个新连接和新节点。
图3:NEAT 的两种结构变异。
1. 添加连接 (Add Connection):在两个节点间增加新连接,赋予新的创新号。
2. 添加节点 (Add Node):在现有连接中间插入新节点。原连接被禁用,产生两个新连接和新节点。
在交叉(Crossover)时,NEAT 利用创新号对齐基因: * 匹配基因 (Matching Genes):创新号相同的基因。随机继承。 * 不相交 (Disjoint) / 过剩 (Excess) 基因:只存在于一方的基因。通常从适应度更高的父代继承。
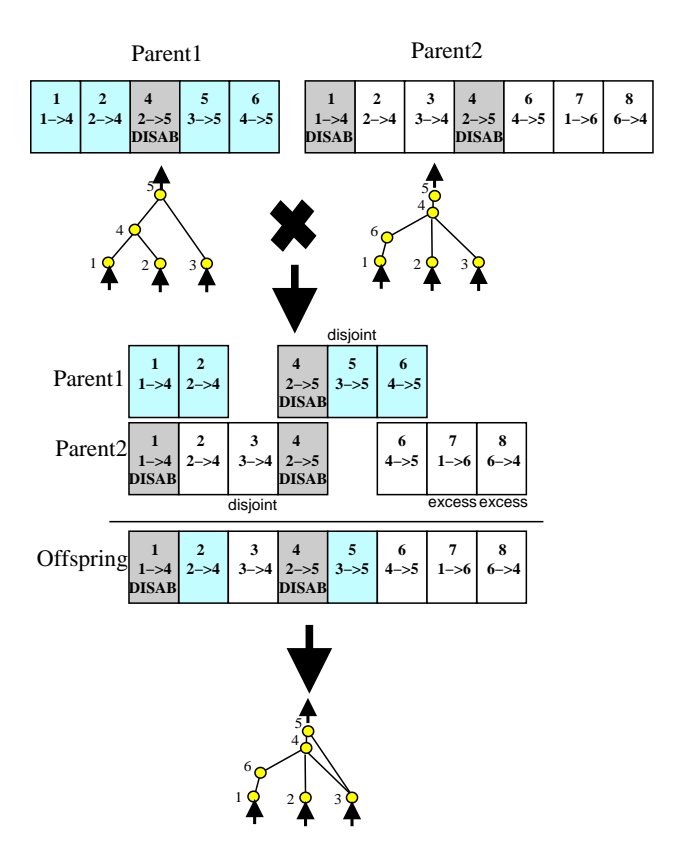 图4:利用创新号匹配不同拓扑结构的基因组。无需复杂的拓扑分析,只需线性对齐创新号即可实现有意义的交叉。
图4:利用创新号匹配不同拓扑结构的基因组。无需复杂的拓扑分析,只需线性对齐创新号即可实现有意义的交叉。
3.3 物种形成 (Speciation) - 保护创新
为了保护创新,NEAT 将种群划分为不同的物种 (Species)。 * 兼容性距离 (Compatibility Distance $\delta$):基于两个基因组中过剩基因 (E)、不相交基因 (D) 的数量以及匹配基因权重的平均差异 ($\bar{W}$) 计算。 $$ \delta = \frac{c_1 E}{N} + \frac{c_2 D}{N} + c_3 \cdot \bar{W} $$ * 显式适应度共享 (Explicit Fitness Sharing):个体必须与同物种的其他成员共享适应度。这限制了任何单一物种过度统治种群,并保护了新产生的小物种(创新),让它们有时间优化自己的结构。
3.4 从最小结构增量增长 (Incremental Growth from Minimal Structure)
与其他通常从随机拓扑开始的方法不同,NEAT 从没有任何隐藏节点的最小网络开始(所有输入直接连接到输出)。 * 随着进化进行,通过变异逐步增加结构。 * 这确保了 NEAT 始终在解决问题所需的最小维度的搜索空间中进行搜索,从而极大提高了效率。
4. 实验验证 (Performance Evaluations)
4.1 XOR 问题 (验证性实验)
XOR 问题需要隐藏层才能解决。 * 结果:NEAT 能够可靠地进化出解决 XOR 问题的结构。 * 效率:平均在 32 代内找到解决方案,且生成的网络结构非常紧凑(通常只有 1-2 个隐藏节点)。
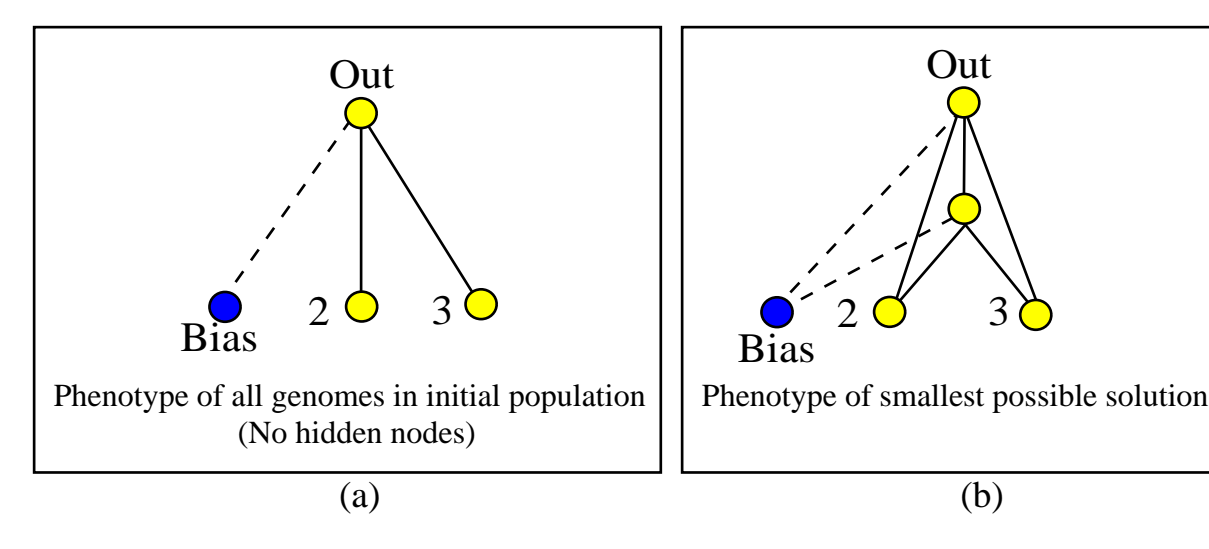
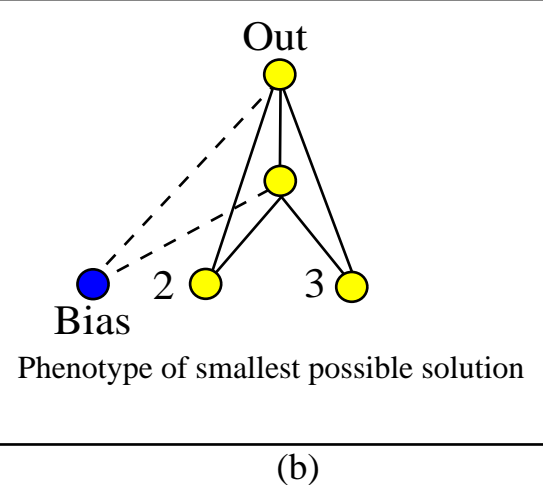 图5:(a) 初始种群没有隐藏节点。(b) 进化出的最优 XOR 网络结构。
图5:(a) 初始种群没有隐藏节点。(b) 进化出的最优 XOR 网络结构。
4.2 双倒立摆平衡 (Double Pole Balancing)
这是一个具有挑战性的强化学习基准任务,要求同时平衡两根长短不一的杆子。
实验 1:带速度信息 (With Velocity)
- 对比算法:Evolutionary Programming, Conventional NE, SANE, ESP。
- 结果:NEAT 所需的评估次数最少,证明了其在标准任务上的高效性。
实验 2:无速度信息 (Without Velocity) - 困难任务
- 这是一个非马尔可夫 (Non-Markovian) 任务,网络必须进化出循环连接 (Recurrent connections) 来拥有记忆功能。
- 对比算法:Cellular Encoding (CE), ESP。
- 结果:
- NEAT 比 CE 快 25倍。
- NEAT 比 ESP 快 5倍。
结论:在困难任务中,进化拓扑结构的优势最为明显。NEAT 能够避免陷入局部最优(Deception)。
5. 分析与消融研究 (Analysis & Ablations)
为了证明 NEAT 的每个部分都是必要的,作者进行了消融实验(移除某个组件后测试性能)。
| 实验设置 | 结果 (平均评估次数) | 失败率 | 说明 |
|---|---|---|---|
| Full NEAT | 3,600 | 0% | 完整版本性能最优 |
| Non-mating NEAT | 5,557 | 0% | 移除交叉:证明了 NEAT 的交叉机制是有效的 |
| Initial Random NEAT | 23,033 | 5% | 随机初始拓扑:证明了从最小结构开始的重要性 |
| Non-speciated NEAT | 25,600 | 25% | 移除物种形成:证明了保护创新的必要性 |
| No-Growth (Fixed) | 30,239 | 80% | 固定拓扑:证明了进化拓扑结构的巨大优势 |
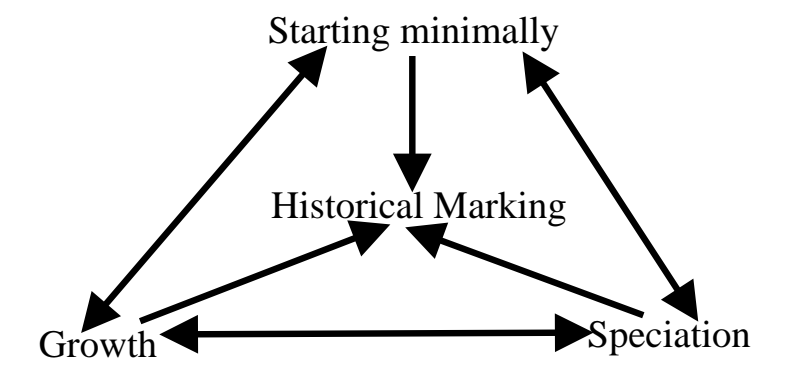 图6:NEAT 各组件之间的依赖关系。历史标记是基础,支持交叉和物种形成;物种形成支持增量增长(保护创新);增量增长确保搜索空间最小化。
图6:NEAT 各组件之间的依赖关系。历史标记是基础,支持交叉和物种形成;物种形成支持增量增长(保护创新);增量增长确保搜索空间最小化。
可视化物种形成
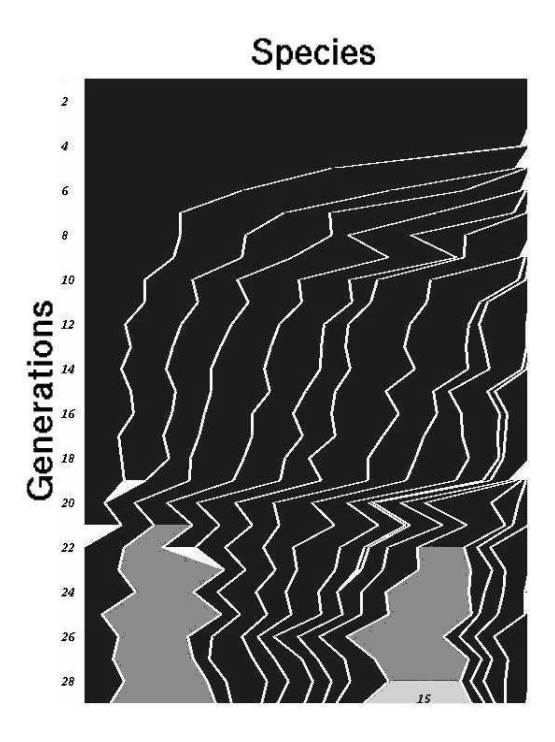 图7:双倒立摆任务中的物种演化图。横轴代表种群大小,纵轴代表代数。可以看到新物种(右侧)不断产生,旧物种(左侧)逐渐灭绝。高亮部分表示该物种内出现了高适应度的个体,最终导向解决方案。
图7:双倒立摆任务中的物种演化图。横轴代表种群大小,纵轴代表代数。可以看到新物种(右侧)不断产生,旧物种(左侧)逐渐灭绝。高亮部分表示该物种内出现了高适应度的个体,最终导向解决方案。
6. 讨论与结论 (Discussion & Conclusion)
NEAT 的优势
- 效率:通过始终在最小维度的空间搜索,NEAT 比固定拓扑方法和其他 TWEANN 更快。
- 复杂化 (Complexification):NEAT 不仅是在优化,而且是在复杂化解决方案。它从简单开始,随着任务需求增加复杂度。这与生物进化的过程高度相似。
- 避免局部最优:通过同时维护不同拓扑结构的物种,NEAT 不容易陷入单一拓扑的局部最优解。
结论
NEAT 证明了进化拓扑结构如果方法得当(使用历史标记解决交叉、使用物种形成保护创新、从最小结构开始),可以显著优于固定拓扑的神经进化方法。它为持续协同进化 (Continuous Coevolution) 和复杂系统的自动生成提供了新的可能性。
 图8:NEAT 为无速度双倒立摆问题找到的一个优雅且紧凑的解决方案。它利用自身的一个循环连接计算导数,从而推断出速度信息。
图8:NEAT 为无速度双倒立摆问题找到的一个优雅且紧凑的解决方案。它利用自身的一个循环连接计算导数,从而推断出速度信息。